 A fundamental, and often false, prediction of evolution is that functionally unconstrained DNA should not be conserved in distant species. As described here, everything from proteins to long stretches of DNA, though highly similar across different species, defy evolutionsts who search for a functional constraint, as required by their theory. And how do evolutionists respond? They claim that the mystery DNA must have a function, even though myriad tests have found none. If we cannot detect the function, then there must be a problem with the tests. There must be a function which otherwise is undetectable to us. It is a perfect example of how religious belief resists falsification as Antony Flew once illustrated in his gardener parable:
A fundamental, and often false, prediction of evolution is that functionally unconstrained DNA should not be conserved in distant species. As described here, everything from proteins to long stretches of DNA, though highly similar across different species, defy evolutionsts who search for a functional constraint, as required by their theory. And how do evolutionists respond? They claim that the mystery DNA must have a function, even though myriad tests have found none. If we cannot detect the function, then there must be a problem with the tests. There must be a function which otherwise is undetectable to us. It is a perfect example of how religious belief resists falsification as Antony Flew once illustrated in his gardener parable:
Once upon a time two explorers came upon a clearing in the jungle. In the clearing were growing many flowers and many weeds. One explorer says, "Some gardener must tend this plot." The other disagrees, "There is no gardener." So they pitch their tents and set a watch. No gardener is ever seen. "But perhaps he is an invisible gardener." So they set up a barbed-wire fence. They electrify it. They patrol with bloodhounds. (For they remember how H. G. Well's The Invisible Man could be both smelt and touched though he could not be seen.) But no shrieks ever suggest that some intruder has received a shock. No movements of the wire ever betray an invisible climber. The bloodhounds never give cry. Yet still the Believer is not convinced. "But there is a gardener, invisible, intangible, insensible, to electric shocks, a gardener who has no scent and makes no sound, a gardener who comes secretly to look after the garden which he loves." At last the Sceptic despairs, "But what remains of your original assertion? Just how does what you call an invisible, intangible, eternally elusive gardener differ from an imaginary gardener or even from no gardener at all?"
Darwin and the evolutionists see a gardener, and to these motivated believers evolution is true despite even glaring question marks and failed expectations that have other observers wondering how evolutionists can be so sure their idea is a fact. And as in Flew's parable, sometimes evolutionists have creative explanations for why the evidence does not seem to reveal their gardener.
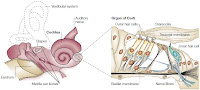
Consider prestin, a protein important in mammalian hearing. Prestin, a transmembrane protein in the outer hair cells of the cochlea, is a frequency-selective amplifier in a sound system that works something like this.
As sound enters the ear, it deflects the outer hair causing tiny amounts of stretching or compression in the outer hair cells. There are channel proteins that sit in the membrane of these cells which are sensitive to such mechanical strain. These proteins provide a tunnel (or channel) across the membrane so that ions can easily cross, and the mechanical strain can cause the channels to open.
These channels are precisely designed to allow only certain types of ions to cross. For example, some channels allow the positively charged potassium ion to cross but not the positively charged sodium ion, and vice-versa.
When a channel opens, ions usually tend to cross through the membrane (either into the cell or out of the cell) because the ion concentration is not uniform, and because there is a voltage, across the membrane. Such differences in concentrations across the membrane, and the voltage, are actively maintained by the cell. They serve as a sort of battery whose energy can be tapped at any time by opening membrane channels.
When the incoming sound causes certain channels to open, the ions that cross cause a change in the membrane voltage. In the outer hair cells, this voltage change encourages negatively charged chlorine ions to exit the cell. They interact with the prestin protein, in the membrane, to cause a mechanical deformation resulting in the elongation of the cell.
In other words, the incoming sound, that caused the hair to move, ends up causing yet more hair movement, and this serves precisely to amplify the incoming sound. This amplification is greater at low sound levels, as it should be.
One of the interesting features of this system is the speed at which it operates. Obviously in order to amplify sound you need to respond as fast as the changes in sound occur. Protein motors often use chemical energy (such as the splitting of the ATP molecule) but that would be too slow for the ear's sound system. Instead, prestin uses the membrane's voltage. This electrical energy can be used much faster and prestin operates at microsecond rates. Here is how
one paper summarized the system:
The exquisitely high sensitivity and frequency selectivity of the mammalian hearing organ originates from a mechanical amplification mechanism that resides in the organ of Corti, the sense organ of hearing in mammals. The gain provided by this amplification can reach as high as a thousandfold; it is highest at low sound levels and progressively diminishes with increasing sound energy.
Of course evolution has no explanation for the origin of this system beyond unfounded speculation, but that is no surprise.
New research is, however, adding a twist to the story.
We now know that prestin proteins in certain bat and whale species (used in their biosonar systems) are more similar than evolution would expect. The massive prestin protein has too many amino acids that match up between these species. If one were to construct an evolutionary tree on the basis of prestin comparisons alone, then the bat and whale would be grouped together, and that cannot be correct.
This fact alone need not be a problem for evolutionists. They simply say that prestin is under the influence of strong selection. In other words, there are strong functional constraints on prestin that require more similarity, even between distant species, than we typically find in proteins.
In particular, researchers identified nine amino acids in prestin that seem to be responsible for the overly-consistent whale-bat matchup. Those nine amino acids must be under very strong selection. If one of them mutated then the biosonar system would not work well. The bat or whale would not survive, and that is why we don't observe such changes. That is how natural selection works.
But if all nine amino acids are required, how did evolution stumble onto the design in the first place? It would be highly unlikely for the right nine amino acids to arise via blind mutations, at the same time.
Of course this conundrum is not confined to the prestin protein. Biology is packed with incredible designs that appear to be finely-tuned. And dozens of finely-tuned design parameters do not appear to be the result of evolution.
The typical evolutionary explanation for biology's wonders is that those design parameters evolved one at a time. It was a gradual process as each setting independently contributed to increased fitness. In other words, a long series of lesser designs preceded today's marvels.
But if a lesser design was good enough before, then why is it not good enough today? Why are those nine amino acids maintained today in prestin if before only a few were required for success?
The evolutionary answer is that there was an arms race. Yes, lesser designs did the job before, but that was then and this is now. The competition has improved and so what worked before no longer does the job.
But like Flew's gardener, these other worlds are forever up for speculation. The competition may or may not have had this or that capability at this or that particular time. The precise timings and interactions can always be tweaked to fit the evolutionary narrative.
And if it turns out that prestin is not so sensitive to some of those nine amino acids, evolutionists won't skip a beat as they didn't before when conserved designs were found to lack the expected sensitivities. One way or another, there is a gardener. Religion drives science, and it matters.
 At the First Church of Darwin, at the University of Chicago, Reverend Jerry Coyne preaches on evolution. Reverend Coyne's preaching is very much in the Darwinian wisdom literature tradition, explaining all manner of religious doctrines that, as evolutionists have explained many times, we all must accept. As a service, Reverend Coyne has gathered those theological dictates he has deemed most important to the Church of Darwin in his book Why Evolution is True. These truths need to be preached to all peoples, and so here we begin. Here is, then, the Word of Coyne:
At the First Church of Darwin, at the University of Chicago, Reverend Jerry Coyne preaches on evolution. Reverend Coyne's preaching is very much in the Darwinian wisdom literature tradition, explaining all manner of religious doctrines that, as evolutionists have explained many times, we all must accept. As a service, Reverend Coyne has gathered those theological dictates he has deemed most important to the Church of Darwin in his book Why Evolution is True. These truths need to be preached to all peoples, and so here we begin. Here is, then, the Word of Coyne:



















